Ingeniørprinsipper forklarer livet bedre enn evolusjonære prinsipper
 Fenotype plastisitet
Fenotype plastisitet
Fenotypisk plastisitet refererer til en organismes evne til å transformere sin anatomi og fysiologi som respons på miljømessige stimuli. Endringene gjør ikke resultat av genetiske endringer, men fra interne adaptive mekanismer. Ralf Sommer oppregnet disse mekanismens essensielle komponenter i en Gjennomgangsartikkel (Sommer 2020): Plastisitet krever utvikling av omprogrammering i form av utviklingsmessige brytere som kan inkorporere miljøinformasjon. Imidlertid de tilknyttede Molekylære mekanismer er kompliserte, og involverer komplekse plasseringer, for eksempel EUD-1, at funksjon som brytere og GRN -er. Selv om det fortsatt er tidlig, er det sannsynlig at på/av-gener peker på et generelt prinsipp om plastisitet, fordi andre eksempler på plastisitet også involverer komplekse brytermekanismer. (s. 7)
Bilde 1. Sopp kan tilpasse seg omgivelene
'Inkorporering av miljøinformasjon' innebærer stilltiende tilstedeværelse av sensorer og signaloverføringsveier. Bryteren som inkluderer sensorisk utgang, tilsvarer en logikkbasert analysator, og genregulerings nettverksaktivitet tilsvarer utgangsresponsen. Kjerne- komponenter samsvarer perfekt med ingeniørmodellen for tilpasning. Fenotypisk plastisitet har blitt observert hos mange arter i forskjellige taxa. Mus utsatt for kaldt vær vil produsere avkom med kortere og færre ryggvirvler (Ballinger & Nachman 2022). I et mer forbløffende eksempel, måker av familien Laridae sporer natriumnivået i blodet, med sensorer i hjertekarene (Sarras et al. 1985). Når nivået når en viss terskel, genererer måker en spesialisert kjertel som trekker ut overflødig natrium fra blodet, og skiller det ut gjennom nebbet. Hvis måken migrerer til et ferskvannsmiljø, forsvinner kjertelen.
Anolis øgler oppvokst på vide overflater, utvikler lange ben som tillater dem større sprinthastigheter, og øgler oppvokst på små overflater utvikler korte ben som gir mer forsiktige bevegelser (Losos et al. 2000). Rove biller svare på tilstedeværelsen av soldatmaur, ved å transformere kroppene, lukten og bevegelser, for å etterligne maurene (Maruyama & Parker 2017). Mange andre eksempler har blitt identifisert i nyere litteratur.
Modellorganismer bekrefter ingeniørbaserte modeller
Mange modellorganismer har dramatisk endret anatomi og fysiologi da de tilpasset seg nye miljøer. Begrepet modellorganisme refererer til en art som er mye studert fordi den er lett å avle og opprettholde i laboratorieinnstillinger. Nøkkelegenskaper av interesse er også tilgjengelige for undersøkelse, som hvordan kroppsform tilpasser seg skiftende miljøer. Standard evolusjonshistorie har vært at trekk endret seg gradvis, på grunn av tilfeldige mutasjoner og naturlig seleksjon. Slike endringer kan potensielt akkumuleres over lengre perioder, som resulterer i storskala evolusjonære transformasjoner. Men eksperimenter og observasjonsstudier over de siste tiårene, endrer fortellingen. Observert variasjon i alle de fleste ikoniske modellorganismer, som opprinnelig ble påstått å støtte evolusjonær teori, er nå kjent for å støtte ingeniørbaserte modeller.
Bilde 2. Bananflue
 Insekt vinger
Insekt vinger
Forskning på Drosophila -fruktfluer viser stramme begrensninger på vingen variasjon. Matematisk biolog Vasyl Alba og teamet hans analyserte bananflue vinge-diversitet (Alba et al. 2021). Forskerne anvendte en sofistikert analytisk metode for å kartlegge forskjellene i vingestruktur mellom individfluer i en enkelt befolkning og mellom forskjellige populasjoner. De fant at den grunnleggende arkitekturen til vingene, aldri endret seg. De forklarte varighet av den underliggende strukturen, som følge av 'globale begrensninger innen utviklingsprogrammet', som sikrer lignende utfall.
Med andre ord, forhindrer utviklingsprosesser vinger fra fundamental endring, når de er forstyrret av mutasjoner og miljøsvingninger. Komplekse mekanismer kompenserer kontinuerlig for slike forstyrrelser, for å holde embryoets utvikling på kurs. Tidligere forskning identifiserte slike korrigerende mekanismer som et sentralt trekk, ved utvikling i kompleks organismer (Pezzulo & Levin 2016).
Noen mutasjoner påvirker utviklingen tilstrekkelig med at korrigerende mekanismer ikke kan opprettholde kjernedesignarkitekturen. Slike 'makromutasjoner' er alltid ødeleggende, om ikke dødelige. De negative konsekvensene bekrefter ytterligere ingeniørprinsippet om at enhver endring av parametere utover forhåndsdefinerte designbegrensninger, resulterer i systemfeil. Denne konklusjonen er bekreftet av det faktum at det ikke eksisterer noen genetisk variasjon i noen art som vil la den endres fundamentalt. Genetikeren John F. McDonald kalte denne observasjonen et 'stort darwinistisk paradoks' (McDonald 1983).
I tillegg koblet forskerne den mindre variasjonen som eksisterte, til 'veldig svake mutasjoner' og til fenotypisk plastisitet. Eksempler på sistnevnte, inkluderer en flue-utvikling med modifiserte vinger, som respons på endret temperatur og kosthold. Bemerkelsesverdig var at teamet var i stand til å kartlegge variasjonen fra begge kilder til forskjeller i verdien av en enkelt sammensatt variabel, som først og fremst tilsvarte de nøyaktige stedene, der vingevener krysser hverandre. Alba et al. (2021) oppsummer arbeidet deres som følger:
Bemerkelsesverdig, finner vi at resultatene av vingeutvikling kan være statistisk beskrevet av en endimensjonal (1D) lineær manifold i et morfologisk rom, som tilsvarer en ikke-intuitiv kombinasjon av strukturelle variasjoner på tvers av vingen. Denne dominerende modusen er systematisk begeistret av genererte varianter av veldig svake mutasjoner i signalveis-gener, så vel som av termisk og kostholds-miljø-forstyrrelser. Som sådan gir vårt arbeid direkte empirisk bevis for tilstedeværelse av globale begrensninger innen utviklings-programmet av vingen, trakterende miljøinnganger og genetisk variasjon i fenotyper strukket langs en enkelt akse i morfologisk rom. (s. 2)
Disse observasjonene samsvarer perfekt med de ingeniørbaserte spådommene som at variasjon eksisterer bare for forhåndsdefinerte, justerbare parametere, og verdiene til beslektede variabler (f.eks. plasseringer av vingevene -kryss) er tett avgrenset. Annen forskning validerer sporingsmodellen, for tilpasning for bananfluer, for å demonstrere at selv variasjonen i de justerbare parametrene gjorde at det ikke stammer fra tilfeldige mutasjoner, men fra konstruerte adaptive mekanismer.
Den genetiske variasjonen ser ut til, i stor grad å ha resultert av NGE. Opptil 80% av 'spontane' mutasjoner i bananfluer initieres av TEs (Transponerbare Elemen ter; Mérel et al. 2020). Weber et al. (2005) identifiserte flere TE -innsettinger som påvirker vingestrukturen. Flere studier har identifisert andre eksempler på fenotypisk plastisitet i insektvinger. For eksempel demonstrerte Bitner-Mathé og Klaczko der fruktfluer reagerer på endret temperatur og populasjonstetthet, ved å endre Deres vingestørrelse, form og venemønster (Bitner-Mathé & Klaczko 1999). Fraimout et al. (2018) beskrev hvordan bananfluer reagerer på lavere temperaturer ved å øke forholdet mellom vingestørrelse og kroppsstørrelse. Reduksjonen i belastningen på vinger kompenserer for kaldindusert lavere muskeleffektivitet. De beskrev også hvordan fluer endrer formen på vingene, for å øke slagkraften for å kompensere for lavere taktfrekvens.
Gao et al. detaljerte et enda mer dramatisk eksempel hos plantehoppere (Gao et al. 2019). Disse insektene sporer miljømessige signaler som temperatur, matkvalitet og populasjons- tetthet. Avhengig av forholdene kan de generere to forskjellige versjoner (aka fenotyper eller morfer). I næringsrik miljøer, nymfer utvikler seg til den 'kortvingede' fenotypen med reduserte eller manglende vinger, redusert flyge-muskelvev og lavere energi-metabolisme. Denne formen er mer effektiv i et lokalt miljø med rikelig mat. Enda viktigere er at denne fenotypen reproduserer tidligere og produserer mer egg, som kommer befolkningen til gode under så gunstige forhold. Den 'langvingede' fenotypen genereres hvis miljøforholdene forverres på grunn av redusert matforsyning eller overpopulasjon . Denne versjonen Har et velutviklet flyve-apparat med lengre vinger, større flymuskulatur og en forbedret energi-metabolisme. Det kan unnslippe det ugunstige miljø og kolonisere nye lokaliteter.
Byttemekanismen mellom fenotypene er svært intrikat. Miljøsensorer fôrer avlesninger til IIS -signalveiene, som ofte er ansatt i komplekse organismer for å kontrollere vevsvekst og kropp størrelse. IIS -signalkomponenten utløser en rekke andre mekanismer som styrer hundrevis av gener, inkludert produksjon av alternative versjoner av proteiner. Oppfinnsomheten og kompleksiteten bak byttemekanismer, til og med de enkleste av dyr (Sieriebriennikov et al. 2018), og den andre refererte til mekanismer som ikke bare fremhever det ekstraordinære designet bak biologisk tilpasning, men de forsterker også behovet for ingeniørmodeller, verktøy og språk for å utvide og utvide vår forståelse av levende systemer.
Bilde 3. Ulike typer siklider
 Siklider
Siklider
Den dramatiske variasjonen i siklidefisk har blitt et plakatbarn for det kreative kraften til evolusjonsprosesser, men forskning de siste 15 årene har vist det motsatte. Cooper et al. demonstrerte at variasjon i siklide foring, er stramt begrenset (Cooper et al. 2010). Etterforskerne brukte en hovedkomponentanalyse (PCA) for å kvantifisere forskjeller i fôring Strukturer for cichlidbestander i innsjøene Tanganyka, Malawi og Victoria.
Denne teknikken skaper sammensatte variabler som fanger opp hvordan verdien av en variabel (f.eks. hodebredde) korrelerer med en annen variabel (f.eks. kjevelengde) eller variabler for hver fisk. Analysen avdekket at variasjon i alle populasjoner i stor grad bosatt langs de samme to komposittvariablene (også kjent som hovedkomponentakser). Kara Feilich utførte en PCA på siklide kropps- og finstrukturer, og hun også fant at den samme variasjonen skjedde gjentok seg flere ganger i separate populasjoner (Feilich 2016):
Kroppsformforskjell blant siklidefiskene er blitt studert mye, gjentatte ganger demonstrerer vanlige diversifiseringsakser på tvers av mange avstamninger, inkludert Tropheines (Wanek og Sturmbauer 2015), geofaginer (Astudilloclavijo et al. 2015) og andre (Clabaut et al. 2007; Muschick et al. 2012). (s. 2260). Feilich bemerket at endringer i trekk var sterkt korrelert, for å optimalisere hydrodynamikk og manøvrering av sentral betydning, den underliggende designens arkitektur endret seg aldri. Bare de fleksible parameteene ble justert til finjusterte individer i en populasjon, for å prestere best mulig i si umiddelbare miljøforhold.
Andre studier identifiserte fenotypisk plastisitet i flere egenskaper. Mazzarella et al. alet opp siklider i vann med forskjellige saltholdighetsnivåer. Større saltholdighet resulterte i at voksen fisk viser mindre kropper og lengre kjever (mazzarella et al. 2015). Härer et al. (2019) utsatte Midas -siklider for lys av forskjellige frekvenser. Som svar på en hyppighetsendring, byttet cichlidene uttrykk for kjegle opsin-gener, som er avgjørende for fargesyn, på bare noen få dager.
Parsons et al. Fed to separate grupper av unge yngelfisk forskjellige dietter (Parsons et al. 2016). De to gruppene utviklet seg til voksne med utpreget ulike hode-kjeve-strukturer, som var skreddersydd for å fôre for den tilgjengelige mat, mest mulig effektivt. Navon et al. rapportert om et lignende eksperiment som bekreftet Parsons et al. Resultater (Navon et al. 2021). Etterforskerne demonstrerte også kostholds-induserte adaptive endringer i kroppsform og strålefinne-antall. Den observerte ulikheten mellom de to kostholds-gruppene etterlignet forskjellene mellom distinkte siklide -arter i naturen.
Forskere erkjenner i økende grad at den mest betydningsfulle variasjonen i siklide-fisk resultater, ikke fra tilfeldige mutasjoner, men fra interne adaptiv mekanismer. Parsons et al. (2016) bemerket: Her er et gryende syn på at additiv genetisk variasjon, utgjør en relativt liten prosentandel av fenotypisk variasjon og snarere er det konteksten der trekk utvikler seg, som bestemmer deres endelige form (Hallgrimsson et al. 2014; Hendrikse et al. 2007; Jamniczky et al. 2010; Pfennig et al. 2010). (s. 6021)
Andre etterforskere oppdaget at noe av den viktigste genetiske variasjonen som gjorde at det eksisterende resulterte fra genetiske endringer som peker på NGE. Carleton et al. demonstrerte at noe av den viktigste variasjonen skyldtes TE-er som var målrettet mot genene, som ligger til grunn for syn (Carleton et al. 2020).
Piggfinnefisk
Som med cichlider, demonstrerer alle studier på piggfinnefiske-mangfold at variasjon er tett begrenset. Hohenlohe et al. dokumenterte hvordan det samme ingeniørprinsippet forklarer biologiske systemer bedre enn evolusjonsteori. Variasjon i trekk og genetikk vises gjentatte ganger i separate populasjoner (Hohenlohe et al. 2010): Genomiske regioner som viser signaturer av både balansering og divergerende utvalg, var bemerkelsesverdig konsistente over flere, uavhengig avledede populasjoner, som indikerer at replikering av parallell fenotypisk evolusjon i piggfinnefisk kan være forekommende gjennom omfattende, parallell genetisk evolusjon i en genom-bred skala. (s. 1)
Miller et al. kom til samme konklusjon etter å ha analysert de genetiske forskjellene mellom populasjoner, som bebodde innsjøer i nærvær og fravær av Prickly Sculpin, en fisk som er et rovdyr for piggfinnefisk (Miller, Roesti & Schluter 2019). Piggfinnefisk som samhandlet med stikkende skulpin, skaffet seg raskt lignende endringer hos hundrevis av de samme genene. De to studiene demonstrerer at piggfinnefisk tilpasser seg genetisk til miljøendringer, forutsigbart og hurtig.
 Disse observasjonene antyder at NGE kan drive målrettet genetisk endringer. En annen studie har identifisert det mer eksplisitt. Ishikawa et al. oppdaget at flere piggfinnefisk arter dupliserte FADS2 -enzymet slik at arten kan syntetisere den essensielle fettsyren DHA (Ishikawa et al. 2019). Denne forbedrede evnen tillot dem å kolonisere DHA-mangelfulle ferskvannsmiljøer. Forfatterne antyder at duplikasjonene var tilrettelagt av NGE, muligens flytting av TES.
Disse observasjonene antyder at NGE kan drive målrettet genetisk endringer. En annen studie har identifisert det mer eksplisitt. Ishikawa et al. oppdaget at flere piggfinnefisk arter dupliserte FADS2 -enzymet slik at arten kan syntetisere den essensielle fettsyren DHA (Ishikawa et al. 2019). Denne forbedrede evnen tillot dem å kolonisere DHA-mangelfulle ferskvannsmiljøer. Forfatterne antyder at duplikasjonene var tilrettelagt av NGE, muligens flytting av TES.
Bilde 4. Gjørs i ordenen piggfinnefisk, er i abbor-familien
Andre etterforskere identifiserte eksempler av fenotypisk plastisitet. McCairns og Bernatchez oppdaget at piggfinnefisk som bodde i ferskvanns og Saltvanns-soner i en stort elvemunning, måler vannets saltholdighet (McCairns & Bernatchez 2010). De bruker denne informasjonen for å regulere optimalt Uttrykk for gener som kontrollerer transporten av saltioner, slik at fisken kan tilpasse seg til saltholdighetsendringer.
Baker et al. demonstrert at hunner med piggfinnefisk sporer internt fysiologisk informasjon som lipidforsyning og leverglykogennivå og miljø-signaler som tilgjengeligheten av mat og befolkningstetthet (Baker et al. 2015). Ulike ledetråder setter i gang justeringer av slik reproduktive parametere, som en tid for avl, egg-størrelse og samhold eller flokk- størrelse. De endringene forbedrer sannsynligheten for populasjonens fortsatt overlevelse. Forskerne oppdaget også at individuelle fisker koordinerer finjusteringen av flere egenskaper for å sikre optimal reproduktiv suksess.
Som et siste eksempel har Tibblin et al. (2020) hevet piggfinnefisk i et akvarium med forskjellig fargebakgrunn. Etterforskerne etterlignet også Tilstedeværelse av rovdyr ved å jage fisk med et dypnett og innføre kjemisk Selskaper etterligner tilstedeværelsen av gjedde, som er et naturlig rovdyr. Både fargen og rovdyrstimuli utløste endringer i ryggfargen som hjalp fisken med å unngå deteksjon.
Hulefisk
Hulefisk demonstrerer også fenotypisk plastisitet for flere egenskaper. Rohner et al. Oppvokst A. meksikanus-embryoer i vann med lav konduktivitet etterligner huleforhold (Rohner et al. 2013). Embryoene utviklet seg til voksne med betydelig mindre øyne. Corral og Aguirre alte opp A. mexicanus i forskjellige temperaturer og forskjellige nivåer av vannturbulens (Corral & Aguirre 2019).
Variantforholdene resulterte i at voksen fisk var forskjellig i vertebralt antall og kroppsfasong. For eksempel viste fisk alt opp i mer turbulent vann, mer strømlinjeformede kropper og utvidede rygg- og analfinnbaser som forbedret deres mobilitet i den miljøtilstanden. Bilandžija et al. alte opp samme arter i mørket, og fisken utviklet mange hulerelaterte egenskaper, som motstand mot sult og endret stoffskifte og hormonnivå (Bilandžija et al. 2020). Fremtidige studier vil sannsynligvis demonstrere flere eksempler av fenotypisk plastisitet og NGE.
Sammenbruddet av evolusjonsikoner
Siklider, piggfinnefisk og hulefisk representerer tre av de mest ikoniske eksemplene for tilpasning som biologer presenterer som bevis for sannsynligheten for evolusjons-prosesser som driver storskala transformasjoner. Likevel viser forskning over de siste tiårene at evolusjonære og adaptive prosesser er begrenset i disse fiskene, som i alle komplekse organismer, til bare mindre endringer til eksisterende egenskaper, eller til tap eller duplisering av en eksisterende struktur.
Noen tilfeldige mutasjoner oppstår som gir fordeler, men nesten alle ødelegger enten gener eller forårsaker modifikasjoner så trivielle at de aldri kunne akkumuleres for å fundamentalt endre noen egenskap. Denne konklusjonen var bekreftet ved en omfattende undersøkelse av teknisk litteratur utført av Michael Behe i sin bok Darwin DEVOLVES: Den nye vitenskapen om DNA som utfordrer evolusjon (Behe 2019).
Behe adresserer spesielt studier av variasjonen i siklide-gener. En Undersøkelse identifiserte aminosyreendringer i et gen som påvirker fargemønstre. De mutante genene så ut til å miste visse evner, så mutasjoner forringet det originale genet. En annen studie identifiserte en mutasjon i et gen som ga litt forbedret lysfølsomhet på visse dybder (Nagai et al. 2011). I dette tilfellet endret bare en enkelt aminosyre, i ett protein seg. I kontrast er nesten alle de mest betydningsfulle tilpasningene, nøye fra kontrollerte prosesser som etterlater lite til blind sjanse.
Ingeniørmodeller og livsmønsteret
Ingeniørbaserte modeller forklarer også bedre likhetsmønster og forskjeller mellom arter i de høyere taksonomiske gruppene (f.eks. Phyla, klasser og ordrer) enn teorien om felles aner (Miller 2018, 2021b). Dataforsker Winston Ewert demonstrerte at fordelingen av samme genfamilier i forskjellige arter passer langt bedre, i det han omtaler som en avhengighetsgraf-modell enn den felles stamfar modellen (Ewert 2018). Hans sentrale konklusjon er at likheter i livet, representerer moduler som var implementert i forskjellige arter for å oppnå lignende mål. Denne generelle konklusjonen har blitt validert av flere forskningslinjer de siste tiårene.
Bilde 5. Fig 7.1. 'Livets tre'
Sammenbruddet av livets tre (58)
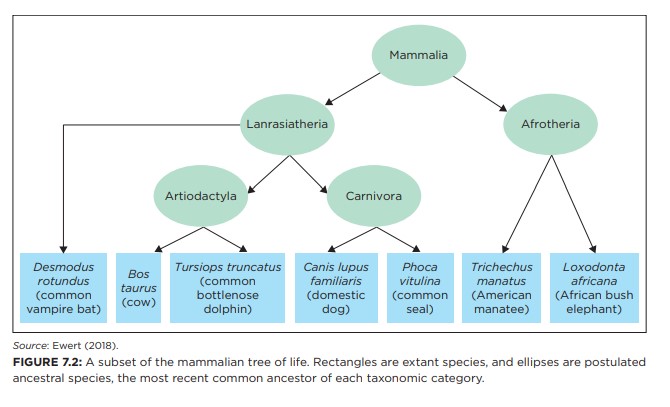
En av de sentrale søylene i standard evolusjonsmodell er troen på a alle levende arter utviklet seg fra en felles stamfar gjennom et gradvis utfoldende livets tre. Som et resultat spår teorien at likhetsmønsteret og forskjeller i arter i dag skal passe inn i et tre-lignende mønster eller nøstet hierarki, der forgreningspunkter tilsvarer utseendet til nye egenskaper. For eksempel deler alle pattedyr visse funksjoner, for eksempel å produsere melk, siden deres siste felles stamfar først utviklet dem. Disse egenskapene ble båret gjennom hver utviklende gren av pattedyrtreet (figur 7.2). Påstanden det alle arter som passer inn i TOL er blitt presentert for publikum som et av de sterkeste bevis for troen på at hele livet utviklet seg gjennom ikke-styrte prosesser.
Bilde 6. Figur 7.2. Pattedyr i livets tre
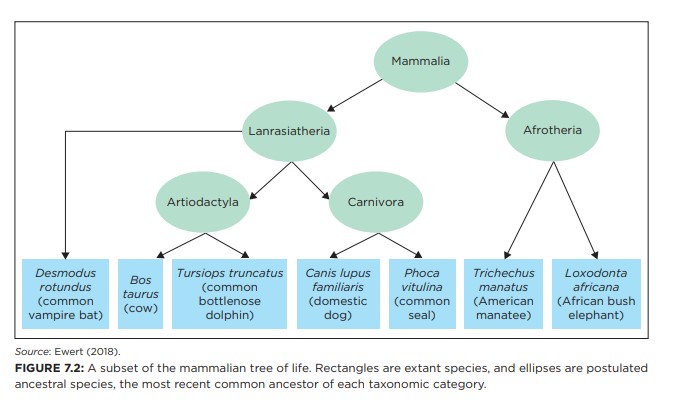
Det offentligheten sjelden blir fortalt er at mønsteret av fysiske egenskaper og molekylære sekvenser i arter, ikke passer inn i en jevnt evolusjonært tre (figur 7.3). Som et ikonisk eksempel har flaggermus og delfiner begge ekkolokalisering, som er undergitt av de samme modifikasjonene på nesten 200 lokasjoner i genomene deres (Parker et al. 2013). Artene antas ikke å være nært beslektet, så likhetene må ha oppstått uavhengig, i direkte konflikt med spådommen om felles aner. Slike konflikter er hyppige og utbredt i nesten alle taxaer.
Zoologer Klassen, Mooi og Locke (1991) oppsummert på tvers av mange, studerer prosentene av egenskaper som passer konsekvent med de best konstruerte evolusjonstrær. Forskerne planla prosentene (også kjent som konsistensindekser) på samme graf som de som er avledet fra tilfeldig genererte data, og verdiene ble deretter justert for å fjerne effekten av tilfeldig støy. Resultatene indikerer at de fleste av dataene ikke passer spådommen om felles aner.
Nyere forskning på omskalerte konsistensindekser (RCI) av mange dyregrupper, bekrefter denne konklusjonen. En RCI på 1 indikerer at alle dataene passer konsekvent med et evolusjonært tre. En RCI på 0 indikerer at Likheter er helt tilfeldige. Data om trekkene i leddyr ga en RCI på 0,39 (Wolfe 2017). RCI for Therapsids (påstått stamfar til pattedyr) var 0,42 (Huttenlocker 2009). RCI for primater og deres påståtte nærmeste pårørende var 0,29 (Bloch et al. 2007). Og RCI for cetaceaner (f.eks. Hvaler og delfiner) og deres påståtte nærmeste slektninger var 0,24 (Thewissen et al. 2001). Evolusjonister siterer disse gruppene som representerer noen av de sterkeste bevisene for felles aner, men deres lave RCI -verdier demonstrere det motsatte.
Bilde 7. Figur 7.3. Funksjonell avhengighetsgraf-fra samme trestruktur
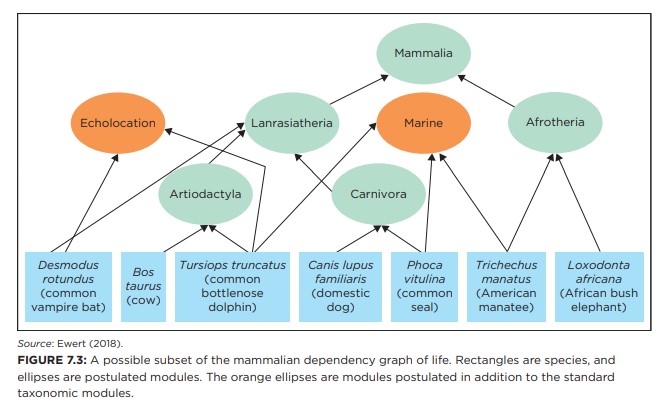
Evolusjonister spådde historisk at RCI -verdier ville falle langt nærmere 1 enn 0, noe de tydeligvis ikke gjør. De lave verdiene viser at antagelse at likheter i trekk, pålitelig innebærer at felles aner er usann.
Problemet har blitt så akutt for mikroorganismer at mikrobiologer Merhej og Raoult beskrev forsøk på å identifisere TOL i de mest negative termer (Merhej & Raoult 2012):
Ingen av de syv poengene som er lagt ut i introduksjonen til dette manuskriptet kan være permanent beholdt, som etablert av Darwins teori, som på den tiden var en kamp mot kreasjonistene. Denne teorien kan ikke opprettholdes i sin helhet. Nyere fremskritt fra genomikk tilbakeviser ideene om gradvis, eksklusiv vertikal arv, evolusjon som velger den mest egnede, en felles stamfar og Livets Tre. Faktisk, er det kanskje ikke engang to gener som har samme evolusjons-treet. (s. 36)
Disse skuffende resultatene har tvunget evolusjonister til å foreslå flere mekanismer for å forklare de tilbakevendende uoverensstemmelsene. Eksempler inkluderer Sideveis GenOverføring (SGO), differensial-gentap, ufullstendig sortering av avstamning og konvergent evolusjon, opp. Sideveis genoverføring refererer til gener som går fra en organisme til en annen. Denne prosessen kan teoretisk forklare hvordan det samme gener vises i ikke -relaterte arter, men sannsynligheten for utbredt SGO i komplekse organismer har blitt stilt alvorlige spørsmål ved (Martin 2017). Ufullstendig avstamnings-sortering og gentap kan ikke forklare komplekse egenskaper som vises i fjernt beslektede organismer. Påstanden om at komplekse tilpasninger kan utvikle seg uavhengig flere ganger (dvs. konvergent evolusjon) kollapser ved nærmere undersøkelse på grunn av usannsynligheten for at de vises gjennom ikke-styrte prosesser, selv en gang.
For eksempel antas det at øyne med linser har utviklet seg uavhengig flere ganger, men alle evolusjonsscenarier står overfor uoverkommelige barrierer. De første mutasjonene i opprinnelsen til et objektiv, ville tildele vev foran fotoreseptorer. Problemet er at udifferensiert vev ville nedbryte Lett mottakelse, så de første mutasjonene ville raskt forsvinne. Objektivet ville ikke bli gunstig før en kompleks utviklingsprosess koblet til et nytt GRN, som dukket opp (Ogino et al. 2012). Likevel den tilgjengelige tiden basert på fossilregisteret utilstrekkelig for selv den minste brøkdelen av nødvendig ny genetisk Informasjon, til å oppstå (Hössjer et al. 2021).
Ingeniørmoduler
I kontrast forklarer ingeniørbaserte modeller tilfredsstillende mønsteret av likheter gjennom hele livet. Designarkitekturer faller ofte inn i et hierarkisk mønster. Alle transportkjøretøyer har visse vanlige funksjoner som for eksempel tildelt plass til last og eller passasjerer, fremdriftssystem og styring. Biler har alle disse funksjonene pluss komponenter som hjul, bremser, kjølevæsker, smøremidler og aksler. Toyota Camry -modeller har alle disse funksjonene pluss ekstra spesialiserte komponenter. Likhetene i transport kjøretøy vil sannsynligvis passe inn i et konstruert tre, i det minste like bra som annerledes grupper av arter.
Mens mange funksjoner i menneskelige produkter passer inn i et hierarkisk tre-lignende mønster, bryter mange det mønsteret. En politibil og et fly har begge toveis-radioer mens toveis radioer er fraværende i de fleste andre biler. i tillegg implementeres samme kretsløp i et bredt utvalg av kjøretøyer for å møte lignende mål. Dette mønsteret gjenspeiler hvordan ingeniører ofte lager moduler som kan være brukt i forskjellige sammenhenger. Modulene må være designet med eksplisitt hensikt å operere i forskjellige produkter, og produktene som bruker moduler må være designet for å innlemme dem riktig i driften. Det samme mønsteret og begrensningene blir observert i livet.
Mange av de samme egenskapene implementeres i forskjellige skapninger å utføre lignende funksjoner. Ulike versjoner av øyne (f.eks. sammensatte og kameralignende) vises uavhengig flere ganger for å tillate optimalt syn i hver art bestemt miljø. Og veldig like nevrale og utviklingsmoduler vises uavhengig av ikke -relaterte arter. Nevro-vitenskapsmenn Sanes og Zipursky identifiserte i både flue og menneskelige visuelle systemer, bemerkelsesverdig lignende designmotiver i netthinnene, nevrale kretsløp i hjernen og genetiske kontrollmekanismer i utvikling (Sanes & Zipursky 2010), selv om de forskjellige øynene til og med antas å ha utviklet seg uavhengig (Piatigorsky 2008).
Tilsvarende demonstrerte matematiske biologer Robyn Araujo og Lance Liotta (2018) at alle biologiske nettverk som utfører 'Robust Perfekt Tilpasning '(RPT) kan deles inn i to forskjellige klasser av moduler. Forskerne definerer RPT AS (Araujo & Liotta 2018):
Evnen til et system til å generere en utgang som går tilbake til et fast referanse nivå (dets 'settpunkt') etter en vedvarende endring i inngangsstimulus, uten behov for innstilling av systemparametere. (s. 2) De samme modulene vises i systemer som er så forskjellige som signaloverføring, genregulering, proteininteraksjonsnettverk, sensoriske systemer og utviklingsmessige regulering. De kombineres ofte hierarkisk for å utføre mer komplekse funksjoner.
Av vital betydning, fungerer basismodulene innenfor krevende begrensninger:
 1. Deres bestanddeler må koble seg sammen i henhold til spesifikke blåkopier.
1. Deres bestanddeler må koble seg sammen i henhold til spesifikke blåkopier.
2. De kan ikke deles inn, så de utgjør et sett med komponenter som er ikke-reduserbart sammensatt.
3. Operasjonene deres må løse en lokal tilpasningsligning, så reaksjoner eller
andre prosesser må operere innenfor snevre grenser.
4. Integrasjonen av modulene i større systemer må også oppfylle rigide
matematiske kriterier.
Bilde 8. Informasjon oppstår ikke av seg selv.
Disse begrensningene innebærer at modulenes opprinnelse og implementering ikke kan ha skjedd trinnvis gjennom en ikke-styrt prosess, idet deres konstruksjon og integrasjon krever koordinering, framsyn og målretting. Szallasi, Stelling & Periwal (2007) antar at evolusjonen må være sant som ett trosengasjement, men de kommenterer fortsatt:
Konseptet med 'modulær design' er lånt fra menneskelig ingeniørvitenskap og har derfor en vesentlig fremtidsrettet, målorientert natur. Komplekse motorer og nettverk er konstruert fra moduler, mens den endelige samlede oppførselen til systemet huskes. (s. 44)
Naturmønsteret tilsvarer også ingeniørprinsipper i distribusjon av artsmangfoldet. I menneskelig ingeniørfag, opprettes mange versjoner av et spesielt design ofte. Hvert år produserer Toyota samme bilmodell med forskjellige sett med moduler som kameraer og sporings systemer. Men få produkter ville falle utenfor de større kategoriene. Kjøretøyer er sjelden designet som vil ligge mellom en ubåt og et fly. {hva design angår -oversetters merknad.}
Tilsvarende eksisterer det mange forskjellige arter som representerer forskjellige versjoner av det samme temaet. Flere ape-lignende og menneskelignende skapninger har vært identifisert, som deler forskjellige likheter. Men likhetene passer ikke innenfor et konsekvent evolusjonært tre (Collard & Wood 2000). Og tydelig mellomliggende skapninger mellom primater og andre ordener av pattedyr, har aldri vært identifisert (Gingerich 2006). Oppsummert peker ikke naturmønsteret mot en ikke-styrt evolusjonsprosess, men ID.
Casestudier på ingeniørmodeller
To casestudier vil ytterligere illustrere ingeniørmodellenes forklarende og forutsigende kraft. Den første fokuserer på minimalt komplekse celler og opprinnelse av livet. Den andre beskriver en ingeniørbasert analyse av flagellum-bakterien.
Minimalt kompleks celle
De to kapitlene av James M. Tour og Fazale Rana detaljerer den usannsynlige av en celle som noen gang har sin opprinnelse gjennom ikke -styrte, naturlige prosesser. Jeg har tidligere forklarte de uoverkommelige termodynamiske utfordringene for en kjemisk system som samles spontant til en autonom celle (Miller 2020a, 2020b). Disse analysene er kritiske for designargumentet. De eneste forklaringer på livet, eller noen gjenstand, er naturlige prosesser, sjanse, sjanse OG naturlige prosesser, eller design (Dembski 2006). Det faktum at naturlige prosesser og eller tilfeldighet ikke kan forklare livets opprinnelse, støtter i seg selv designslutningen. Tillegg av følgende positive bevis gjør identifisering av design avgjørende.
Ingeniøranalyser belyser i selv de enkleste av celler mye av underliggende arkitektur og den klare designlogikken, ovenfra og ned. Top-down logikk krever nivåer av framsyn, koordinering og målretting, som bare intelligente agenter kan implementere. Slike bevis har tvunget til og med forskere som filosofisk avvise design, til ubevisst å anerkjenne det i livets opprinnelses nødvendighet.
Systemingeniører har analysert de minimale kravene til selvutvikling i slike systemer som månefabrikker (Freitas 1980) og roboter (Moses & Chirikjian 2020). Forskningen ble styrt av det teoretiske arbeidet til matematikeren John von Neumann (1966). Parallelt har flere studier forsøkte å identifisere de minimale komponentene i en autonom celle (Xavier, Patil & Rocha 2014). Sammenløp av disse to forskningsstrømmene demonstrerer at en minimalt kompleks autonom celle minst må ha følgende komponenter:
1. Maskiner for energiproduksjon og levering.
2. Informasjonslager og prosessorer.
3. Selektive porter med aktiv transport.
4. Sensorer koblet for å signalisere transduksjonsveier og signalbehandling.
5. Aktuatorer som implementerer instruksjoner.
6. Produksjons- og automatiske monteringsprosesser.
7. Automatiserte reparasjonsmaskiner.
8. Feilkorrigerings-systemer.
9. Avfallshåndtering og gjenvinningsmekanismer.
10. Kontrollsystemer som er i stand til global koordinering.
Teoretiske ingeniøranalyser samsvarer ikke bare med mange av komponentene til minimalt komplekse celler, men også i deres sammenheng og underliggende designlogikk.
 Denne korrespondansen demonstrerer at celler viser top-down design Hvor en overordnet designarkitektur bestemmer systemene på høyere nivå og deres gjensidige avhengigheter. Hvert system fungerer også i henhold til sitt eget designmønster (f.eks. Kontroll tilbakemeldingssløyfe) som bestemmer laveste nivå prosesser. Designmønstrene er demontert fra og overskrider fysikk og kjemi av de underliggende molekylene, så lite som oppstår i en cellw ville skje uten ekstern styring. Ledende opprinnelsesforskere Elbert Branscomb og Michael Russell forklarer hvordan celler må bruke molekylære maskiner for å håndtere slike prosesser som kjemiske reaksjoner (Branscomb & Russell 2018a)
Denne korrespondansen demonstrerer at celler viser top-down design Hvor en overordnet designarkitektur bestemmer systemene på høyere nivå og deres gjensidige avhengigheter. Hvert system fungerer også i henhold til sitt eget designmønster (f.eks. Kontroll tilbakemeldingssløyfe) som bestemmer laveste nivå prosesser. Designmønstrene er demontert fra og overskrider fysikk og kjemi av de underliggende molekylene, så lite som oppstår i en cellw ville skje uten ekstern styring. Ledende opprinnelsesforskere Elbert Branscomb og Michael Russell forklarer hvordan celler må bruke molekylære maskiner for å håndtere slike prosesser som kjemiske reaksjoner (Branscomb & Russell 2018a)
Men selv de av livets molekylære transformasjoner som løper nedoverbakke må bli tatt ut av kjemiens hender og 'administrert' av en dedikert makromolekylær maskin - for å pålegge betinget manipulerbar kontroll over reaksjonshastigheter og å utelukke uønskede reaksjoner, både med hensyn til reaktanter og produkter. I seg selv, er egen kjemi er altfor ukritisk og ukontrollerbar .. (s. 1)
De erkjenner til og med at alt må samsvare med et 'forseggjort organisasjonsdesign'.
Bilde 9. Molekylær maskin (Otangelo Grasso)
Forskerne oppgir at molekylære maskiner ikke bare er nødvendige i livet i dag, men de var også essensielle når det gjelder oppdukking (Branscomb & Russell 2018b, understreking og kursiv -ved oversetter). Bemerkelsesverdig erkjenner de også at selv de mest grunnleggende elementene av livet aldri kunne ha oppstått gjennom naturlige kjemiske prosesser: Vi hevder spesielt at det er uholdbart å hevde at den livsrelevante biokjemien kunne ha dukket opp i det kjemiske kaoset, produsert av masseaksjonskjemi og kjemisk ikke -spesifikke 'energi' innganger, og har først senere utviklet skremmende spesifikke mekanismer (som en del av å utvikle alle resten av livets funksjoner). (s. 4)
De reagerer på denne tilsynelatende uoverkommelige barrieren ved å appellere til naturlig utvalg som en ledende hånd for å bringe hvert system til eksistens og ordentlig forbinde dem. Det åpenbare problemet er at naturlig utvalg ikke kunne ha startet, før en autonom selvrepliserende enhet dukket opp. Forfatterne erkjenner faktisk stilltiende, at livet er produktet av ID, men de bruker språket til naturlig utvalg for å smugle intelligent byrå inn i sin teori, uten å gi behørig kreditt til designeren.
Bakteriell flagellum
Bakteriell flagellum fungerer som et roterende fremdriftssystem i bakterier (Figur 7.4) og viser frem den prediktive kraften til ingeniørmodeller (Miller 2022). Systemet bruker mange fantastisk komplekse delsystemer. Monterings-operasjonen er regissert av et genetisk nettverk (Fitzgerald, Bonocora & Wade 2014) som sikrer fremstilling av riktige proteiner i rette mengder til akkurat rett tid. Proteinene som komponerer propellen er transportert over cellemembranen gjennom en transportport, som bare tillater de korrekte gjennom til riktig tid, ved å bruke protein signalsekvenser (Evans, Hughes & Fraser 2014).
Andre proteiner drifter kroken og propellen (Kim et al. 2017; Ohnishi et al. 1994). Et annet protein fungerer som en linjal som koordinerer med porten, forsikrer deg om at den flagellære kroken strekker seg til riktig lengde (Waters, O'Toole & Ryan 2007). Et navigasjonskontrollsystem kontrollerer rotasjonen til rotasjonsmotoren Motor (Hamadeh et al. 2011).
Dataforsker og ingeniør Waldean Schulz nærmet seg designet av flagellum med en metode som kan beskrives som banebrytende. Han skisserte først hva han forventet ville være den mest effektive arkitekturen for et nano-teknologisk roterende fremdriftssystem (Schulz 2021A). Han startet med det høyeste nivået i det organisatoriske hierarkiet og kartla overordnet design av kjerneprosessene, inkludert produksjon, montering, energiproduksjon, momentgenerering, miljøsporing og retningsbestemt kontroll. Han tegnet de forventede minimale komponentene og deres funksjonelle forhold. Han forventet også designkrav og begrensninger.
Bilde 10. Fig 7.4. Prinsippskisse av bakterieflagellen
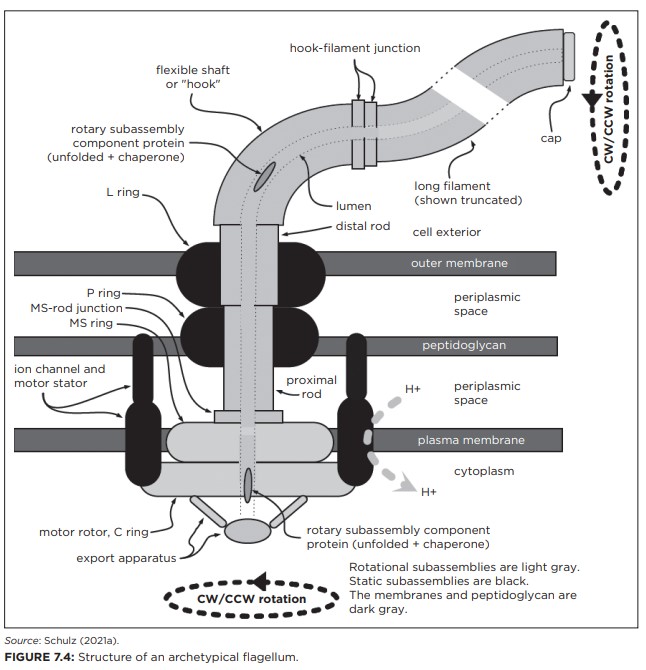
Han gjennomgikk deretter litteraturen om faktiske flagellære operasjoner som starter med prosessene på det nederste organisasjonsnivå (Schulz 2021b). Til slutt, sammenlignet han topp-down og bottom-up-analysene (Schulz 2021c). Schulz's Forventninger til designarkitektur, innbyrdes forhold og begrensninger matcher de faktiske operasjonene i bemerkelsesverdig grad.
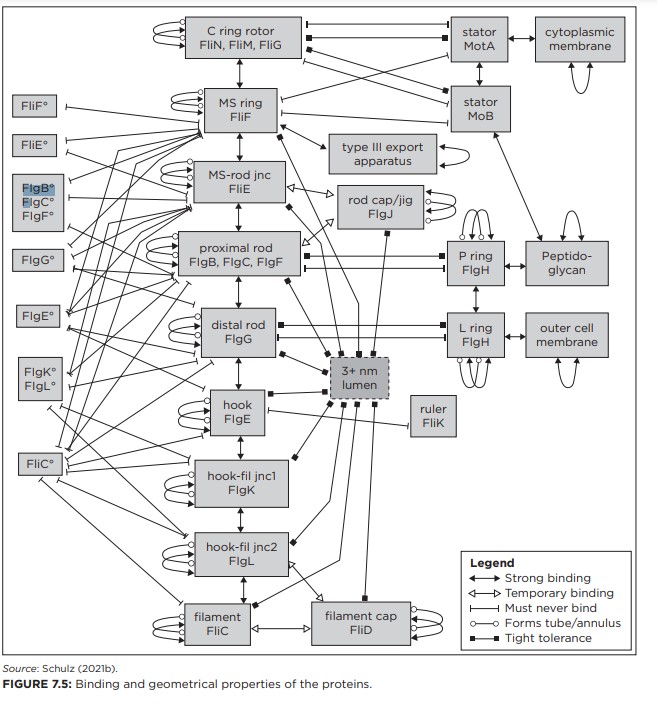
I tillegg identifiserte han mange trange begrensninger som må oppfylles for flagellum for å fungere med en effektivitet som vil gi noen fordel for cellen. Begrensningene inkluderer over 80 krav til samspill mellom de individuelle proteinene (se figur 7.5). For eksempel må noen proteiner binde seg sammen permanent, andre midlertidig, andre må aldri binde seg til hverandre. Hvis noen Av disse kriteriene er ikke oppfylt, vil flagellum ikke gi noen fordel for cellen. Det vil i stedet gi cellen ulemper, ved å kaste bort ressurser.
Schulzs analyse viser at flagellum ble konstruert rundt en klar overordnet designlogikk, der hvert system og komponent var optimalt designet for å integreres med flere andre systemer og komponenter i symfonisk harmoni. Hvert av disse systemene er påkrevd for riktig drift, og de må koordinere med hverandre med ekstremt høy effektivitet. Antall viktige komponenter, innbyrdes forhold, krav og begrensninger i flagellum beviser at systemet må ha sin opprinnelse samtidig, gjennom handlingen til en intelligent agent. Schulzs arbeid er ganske enkelt en forsmak av den langsiktige klarheten og innsikten ingeniørbaserte modeller vil bringe til studiet av levende systemer.
Fremtidig forskning
Fremtidige forskere kan følge Schulzs eksempel ved å identifisere ingeniørfaglig logikk, som ligger til grunn for andre levende systemer. De kan da også forutse kjerne komponenter, deres innbyrdes forhold og de operasjonelle begrensningene av sammenligning av systemene, med sammenlignbare menneskelige kreasjoner. Denne oppgaven er bare mulig fordi livet ikke bruker designmønstre og prosesser som er analoge med de som brukes i menneskelig ingeniørfag. Men de er de samme mønstrene og prosesser.
Livet bruker ikke systemer analogt med informasjonsbehandling, men de er synonyme med informasjonsbehandling. Og celler replikerer ikke DNA ved bruk av systemer, analogt med feilretting, men de utfører feilkorrigering. Denne anerkjennelsen vil unngå villedning, som ofte oppstår fra de falske forut-setningene knyttet til vitenskapelig materialisme (f.eks. Suboptimalt Bunnen-Opp design), og det vil fremskynde fremgangen i biologisk forskning.
Fig 7.5. Analyse av bakterie-flagellen (Schulse)
Konklusjon
Biologifeltet har nettopp begynt å gå inn i den neste store vitenskapelige revolusjonen, der den filosofiske pendelen har begynt å svinge seg bort fra vitenskapelig materialisme og tilbake mot de designbaserte filosofiske tradisjonene til Platon og Aristoteles. Bevisens store tyngde har tvunget fram dette skiftet i det vitenskapelige samfunnet. Spesielt har de sentrale spådommene til evolusjonsteori konsekvent mislyktes, mens spådommene til ID stadig har blitt mer bekreftet. Dette paradigmeskiftet er tydeligst der forskere har de fleste innarbeidede ingeniørprinsipper og innsikt i sin biologiske forskning.
Inkorporering av designkonsepter: Biologer har bare nå begynt å trykke inn i ingeniørmodellers forklarende og prediktiv kraft. Schulz's Anvendelse av ingeniørprinsipper for å analysere flagellum tillot ham å forutse mye av det roterende systemets struktur og operasjoner. Som hans Top-down integrert med bottom-up tilnærming, brukes på andre systemer, Forskere vil i økende grad anerkjenne at de eneste levedyktige rammene for å forstå livet, starter fra antagelsen om design. Denne trenden vil bare fortsette akselerere, i det funn fortsetter å demonstrere ovenfra og ned-naturen av biologisk design, optimaliteten til levende systemer og de stramme iboende begrensningene for biologiske systemer. Mange systembiologer har allerede anerkjent at de må innlemme kjerne -ID -konseptene i analysene sine, om enn ved bruk av forskjellige språk, for å fremme forskningen. Heuristikken for ID er ganske enkelt en mer generell rubrikk, for anvendelse av ingeniørprinsipper på studiet av livet.
Bilde 11. Fig. 7-5 Analyse av bakterie-flagell
Mer spesifikt er Behes begrep om ikke-reduserbar kompleksitet implisitt i holisme-synet, som beskrevet tidligere. William Dembskis formulering av spesifisert kompleksitet, er en mer generell kategori for biologers forståelse av design moduler. Argumentet for design: Dette siste eksemplet er av kritisk betydning. Spesifisert kompleksitet innebærer et mønster, som ikke er et produkt av et naturlig prosess, ekstremt sjelden og samsvarer med et spesielt eksternt definert mønster (Dembski 2006). Mønstre som viser spesifisert kompleksitet utover en viss terskel er alltid produktet av design. Et eksempel ville være et scrabble brev som er arrangert for å stave ut et forståelig avsnitt. Livet demonstrerer mange eksempler på spesifisert kompleksitet; Derfor har Dembski hevdet konklusjonen at livet er designet, er nesten sikkert.
Likevel utfordret evolusjonister Dembski ved å imøtegå, man kan ikke konkluderende demonstrere , at et mønster i biologi på noen måte er spesielt. De hevdet også at en biologisk enhet kan virke svært usannsynlig å ha sin opprinnelse ved en tilfeldighet, men mange andre biologiske løsninger kan også ha spilt den nødvendige rollen. Sannsynligheten for å finne en løsning ut av mangler, kan være gjennomførbar.
Observasjonen at livet benytter de samme designmotiver som er sett i menneskelige Ingeniørfag som moduler, velter disse innvendingene. Et designmotiv er et høyt usannsynlig, spesielt mønster. Relativt få eksisterer i biologi som løser ingeniøroppgaver som RPA. I tillegg det faktum at de samme motivene som ble brukt I menneskelig ingeniørarbeid implementeres gjentatte ganger i livet. Mange andre alternativer eksisterer ikke, som livet kunne ha snublet over ved tilfeldighet.
Bilde 12. Logisk slutning
 De teologiske implikasjonene: denne konklusjonen og tilsvarende paradigmeskifte i biologi, innebærer betydelige teologiske implikasjoner. Mange kristne har forsøkt å forene evolusjonsteori med kristen teologi, men deres innsats har ofte resultert i å besøke eller forvrenge historiske kristne læresetninger og vri bibelske tekster. Eller de har fremstilt evolusjonsteori i termer som direkte er i konflikt med hvordan teorien generelt er forstått og undervist (Moreland et al. 2017). Nå er det vitenskapelige bevis så avgjørende til fordel for design, at kristne ikke lenger trenger kaste bort tid i den meningsløse oppgaven med å la kristen teologi imøtekomme sekulære skapelsesfortellinger.
De teologiske implikasjonene: denne konklusjonen og tilsvarende paradigmeskifte i biologi, innebærer betydelige teologiske implikasjoner. Mange kristne har forsøkt å forene evolusjonsteori med kristen teologi, men deres innsats har ofte resultert i å besøke eller forvrenge historiske kristne læresetninger og vri bibelske tekster. Eller de har fremstilt evolusjonsteori i termer som direkte er i konflikt med hvordan teorien generelt er forstått og undervist (Moreland et al. 2017). Nå er det vitenskapelige bevis så avgjørende til fordel for design, at kristne ikke lenger trenger kaste bort tid i den meningsløse oppgaven med å la kristen teologi imøtekomme sekulære skapelsesfortellinger.
I tillegg demonstrerer designmønstrene som er sett i livet, at Gud skapte verden slik at spesifikke ingeniørmotiver fungerer effektivt for å oppnå slike mål, som syn eller bevegelse. Denne erkjennelsen forklarer det påfallende likheter mellom menneskelige kreasjoner og levende systemer, og det forklarer hvordan Schulz var i stand til å forutsi så mange detaljer om flagellumets struktur og operasjoner.
Kristne kan nå snakke med full tillit til det klare bevis for Guds hånd i livet. Mange ateister og agnostikere som hører sannheten vil fortsette å undertrykke det, men for andre vil bevisene sette tankene fri, for sekulære filosofier. De vil da få frihet til å forfølge et forhold til sin skaper.